
GrainGenes
A Database for Triticeae and Avena
Finch & Bennett, pp. 22-26
II. 6. Variation in expression of tri and somatic instability in P-4 barley.
R. A. Finch and M. D. Bennett, Plant Breeding Institute, Trumpington, Cambridge, England. "R"
The tri gene has great potential for producing easily large quantities of cytological variants and providing material for the study of somatic chromosome elimination. In diploid tri homozygotes of Paavo barley line P-4, 50% of seeds are thin and have triploid embryos (Ahokas, 1977). The tri gene suppresses second meiotic division in 50% of embryo sac mother cells so that they form diploid embryo sacs, but does not otherwise affect ovule development and meiosis is regular in over 80% of pollen mother cells (Finch and Bennett, 1979). In a sample of 277 cells, over 90% of pollen grains showed n=7 at first mitosis. Diploid tri homozygotes grown in favourable conditions can be as fertile on open pollination as Paavo, but ranged from 40% to 90% in fertility in recent tests.
Production of cytological variants
Large numbers of P-4 plants at six ploidy levels have been obtained.
Haploids, together with diploids, were easily obtained by embryo culture
from diploids crossed with 2x Hordeum bulbosum (Finch and Bennett,
1979). To obtain tetraploids, the roots of potted juvenile diploids were
washed free of soil and cut back to 3 cm and plants were immersed in aqueous
colchicine (0.05% w/v) and dimethyl sulfoxide (1.5% v/v) for 5 h at 20°C
under "cook white" fluorescent tubes, repotted and grown in a glasshouse
to maturity. Of 433 fat seeds screened, 11 gave tetraploid plants and 3
gave diploids with 2x/4x mosaic roots. A tetraploid arose spontaneously
on an otherwise typical diploid grown from seed produced by a colchicined
haploid and a low frequency of other anomalous progenies arose from different
treated and untreated P-4 sources (Table 1). In tetraploids, nearly all
fat seeds tested had 4x embryos and nearly all thin seeds tested had 6x
embryos. No systematic search was made, but there is little evidence in
2x, 3x and 4x plants of the high frequency of instability in somatic chromosome
number reported later in 6x plants.
Table 1. Frequencies and types of anomalous progenies from different P-4 sources.
{kind=link}
Fertility on open pollination was nil in haploids, below 1% in triploids and often below 40% in tetraploids. When pollinated by pollen from diploids, fertility in triploids was nearly 3% and chromosome numbers in 30 progeny were 2n=14 (4), 2n-14+1 telocentric (1), 2n=15 (13), 2n=16 (4), 2n=17 (1), 2n=3x (4) and 2n=4x (3). Eleven trisomic plants (2n=15) set no seed on open pollination, 5 set 1-18 seeds each and 2 set well over 100 each. A telotrisomic plant set 18 seeds but 2 plants with 2n=16 and 1 with 2n=17 set no seeds.
Variation in percentage of thin seeds
P-4 plants differed significantly (P < P.001) in the percentage
of seeds that were thin at both 2x (20 plants) and 4x (9 plants) levels,
ranging from 26.2 to 59.5 (mean 45.9 + 2.3) and 46.4 to 73.1 (mean
63.3 + 2.6), respectively. It is unknown if these differences were
genetic, but crosses of 2x P-4 as female with barley line BGM64 (cross
1) and as male with Vada (cross 2) and Betzes (cross 3) gave evidence of
genetic variation among F2 plants in the percentage of thin seeds. The
percentage of thin seeds in different tri plants ranged from 1.6
to 40.0, 1.3 to 54.8 and 1.0 to 51.9 in crosses 1, 2 and 3, respectively,
and 73.3, 60.0 and 64.0% of tri plants had under 10% of thin seeds.
The great variation among plants in percentage of thin seeds may be partly environmental, but in the 3 F2 populations must also be partly genetic. It offers the hope of manipulating genotypes and environments to raise at will the proportion of thin, i.e. polyploid seed. Most F2 segregants had relatively fewer thin seeds than P-4 but this may not be inevitable. Thus in cross 1, F2 plant number 79/95, which was 100% fertile, with 2.1% of thin seeds, gave 51 F3 plants with a mean of 10.5 + 1.3% of thin seeds including one plant wtth 46.2%. If a cross could be made to raise the percentage of thin seeds similarly by over 40% in P-4, very high yields of polyploids might be easily obtained.
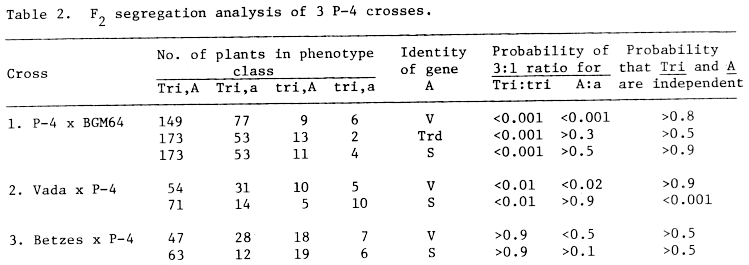
Table 2. F2 segregation analysis of 3 P-4 crosses.
{kind=link}
Segregation of tri in crosses
In each cross, F2 plants were grown together in the same glasshouse
and all plants with 1 or more 3x seeds were scored as tri. Some
tri plants may have been misclassified as Tri since sterility
reached 40.0% (mean 5.1 + 1.8%) 62.5% (mean 24.9 + 2.0%)
and 90.0% (mean 11.8 + 1.9%) in individual plants scored as Tri
in crosses 1, 2 and 3, respectively, and some of the sterile florets might
have had triploid seeds if they had been fertile. Marker genes segregating
with tri were V and S in crosses 1-3 and trd
in cross 1. Table 2 gives linkage data between these genes and tri.
Tri showed a monogenic F2 ratio (cf. Ahokas, 1977) in cross 3, but
dihybrid ratios 15:1 (P > 0.9) and 13:3 (P > 0.5), in crosses 1 and 2,
respectively. The apparent dihybrid ratios are probably due to misclassification,
but it is possible that the tri phenotype may be governed by two
loci. Trd and S showed monogenic ratios, but V consistently
showed a ratio of about 13:7. Cross 2, but not cross 3, indicated linkage
between Tri and S, but this apparent linkage clearly needs
confirmation.
Somatic instability in 5x and 6x plant
About 97% of thin seeds on open pollinated 4x P-4 plants had 6x embryos
and most of the rest had 5x embryos and such seeds are a convenient source
of highly polyploid Hordeum vulgare (Finch, 1980). However, despite
culture in hydroponics (cf. methods in Finch and Bennett, 1981) and in
soil in a variety of glasshouse and growth room conditions, no 6x or 5x
plant survived for more than 29 weeks and most stayed as grass dwarfs like
the six 6x plants of Rommel (1960). Of over 90 plants with 6x roots, only
3 produced ears and in one plant, the ear was fixed before meiosis but
no mitoses were found. Of 16, initially 6x plants sampled twice or more
in separate weeks, 6 showed within-plant variability in chromosome number
(Table 3). Most roots had only 1 chromosome number but 2 or 3 numbers coexisted
in roots of plants 4, 5 and 6 (Table 3). Chromosomes were counted in up
to 15 and up to 24 cells in each root of non-mosaic and mosaic plants,
respectively, but in most roots only 2 or 3 cells were sampled.
In 9 of the 16 plants sampled, the chromosome number in the earliest roots was the same as that in roots sampled 7-29 weeks after germination. In both spikes where chromosome counts were possible, however, the chromosome number was considerably below the 6x level found 15-17 weeks earlier in roots. It is therefore highly probable that chromosome instability is common in hexaploid shoots. The cause of the instability in 6x P-4 is unknown but the phenomenon clearly deserves further study, particularly in comparison with the preferential loss of one parent genome in interspecific hybrids of H. vulgare with other Hordeum species, where spike primordia have a higher elimination rate (i.e. frequency) than roots (Noda and Kasha, 1981).
{kind=link}
References:
Ahokas, H. 1977. A mutant of barley:triploid inducer. Barley Genet. Newsl. 7:4-6.
Finch, R. A. 1980. New source of aneuploids and high polyploids in barley (Hordeum vulgare L.). Heredity 44:285 (Abstr.).
Finch, R. A. and M. D. Bennett. 1979. Action of triploid inducer (tri) on meiosis in barley (Hordeum vulgare L.). Heredity 43:87-93.
Finch, R. A., J. B. Smith and M. D. Bennett. 1981. Hordeum and Secale mitotic genomes lie apart in a hybrid. J. Cell Sci. 52:391-403.
Noda, K. and K. J. Kasha. 1981. Chromosome elimination in triploid hybrids between Hordeum vulgare (2x) and H. bulbosum (4x). Cereal Res. Communs 9:85-91.
Rommel, M. 1960. The occurrence of euploid and aneuploid hexaploid plants within the offspring of artificially induced tetraploid Hordeum vulgare L. Can. J. Genet. Cytol. 2:199-200.