
GrainGenes
A Database for Triticeae and Avena

Are powdery mildew resistance loci Ml-p and ml-d on barley chromosome 5?
Jens JensenAgricultural Department
Risø National Laboratory
DK-4000 Roskilde, Denmark
The loci Ml-p and ml-d were reported linked with locus at (albino seedling) on chromosome 5 with, respectively, 36.65 and 12.08% recombination (Briggs, 1945). Furthermore, the recombination frequency between ml-d and Ml-p was 16.38% (Briggs, 1945), and that between ml-d and Ml-a was 17.5% (Schaller and Briggs, 1955) whereas that between loci Ml-p and Ml-a was very high 41.5 + 2.5%. But, in calculation of the recombination frequencies, the authors paid no attention to possible deviations from the expected Mendelian segregation ratio of each of the powdery mildew resistance genes. If deviation had occurred, the recombination percentages reported may be far from the true ones. However, on accepting the recombination percentages reported (cf. Table 1) it was possible to map loci Ml-p and ml-d on barley chromosome 5 (Jensen and Jørgensen, 1975).
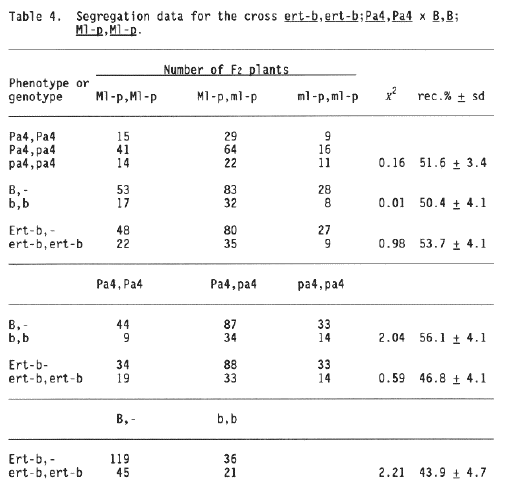
In Barley Genetics Newsletter Vol. 17, locus Ml-p was removed from the linkage map of chromosome 5 (Jensen, 1987) based on some unpublished data. The present report describes these data, which originate from six crosses (Tables 2 to 7). The segregation data from Ml-p combined with translocation T2-5a (Table 2) show no sign of linkage. This was expected if Ml-p is located on chromosome 5 on its most likely position according to the above data (Jensen, 1986).
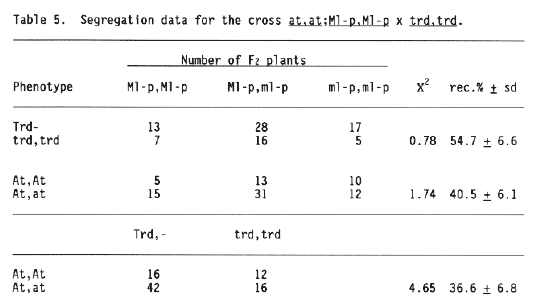
The segregation data in Tables 3 to 7 show no significant linkage to any of the following seven loci, with the map positions spread all over the recent map of chromosome 5 (Jensen 1988): trd (pos. -73 cM) Tables 5 and 6; fl (pos. -58 cM) Tables 3 and 4; at (pos. -14 cM) Tables 5 and 7; nec1 (pos. 0 cM) Table 7; ert-b (pos. 15 cM) Table 4; fs2 (pos. 18 cM) Table 7; and Pa4 (pos. 85 cM) Table 4. The numbers of F2 progenies tested from these crosses are so high that it is most unlikely that locus Ml-p can be located in between any of these seven loci. In addition, it should be noted that locus Ml-p is independently inherited from two other chromosome 5 loci, Ml-k (pos. 63 cM) and Ml-at (pos. 85 cM), and it may be loosely linked, if linked at all, to Ml-a (Table 1). Therefore, it must be concluded that locus Ml-p is not located on chromosome 5.
With respect to the possible association of locus ml-d with chromosome 5 the following considerations are made. Firstly, a consequence of the above is that the linkage reported between ml-d and Ml-p (Table 1) cannot be used to show that ml-d is on chromosome 5. Secondly, the linkage of ml-d with locus at, in position -14 cM on chromosome 5, has a very high standard deviation (Table 1) reducing severely the validity of this estimate. Thirdly, it is incomprehensible that ml-d is linked with Ml-a, but not with Ml-k (Table 1), since it is known that Ml-a and Ml-k are linked with about 8% recombination. Lastly, the recombination reported in Table I may be confounded by possible deviation from the expected Mendelian segregation ratios.




Taken together, the available data give no support for locus ml-d being located on chromosome 5.
The conclusion is therefore: Ml-p is not located on chromosome 5 and it is now more likely that ml-d is located on chromosome 5 than on any of the other barley chromosomes.
References:
Briggs, F.N. 1945. Linkage relations of factors for resistance to mildew in barley. Genetics 30:115-118.
Jensen, J. 1987. Coordinator's report: Chromosome 5. BGN 17:111-113.
Jensen, J. 1988. Coordinator's report: Chromosome 5. BGN 18:61-63.
Jensen, J., and J.H. Jørgensen. 1975. The barley chromosome 5 linkage map. I. Literature survey and map estimation procedure. Hereditas 80:5-16.
Moseman, J.G., and CA. Schaller. 1960. Genetics of the allelic series at the Ml-a locus in barley and cultures of ' Erysiphe graminis f.s. hordei that differentiate these alleles. Phytopathology 50:736-741.
Schaller, CA., and F.N. Briggs. 1955. Inheritance of resistance to mildew, Erysiphe graminis hordei, in the barley variety, Black Russian. Genetics 40:421-428.
Scholz, F., and I. Nover. 1967. Genetische Untersuchungen mit einer vollstandig mehltauresistenten Gerstenlinie. Kulturpflanze 15:242-254.
