
GrainGenes
A Database for Triticeae and Avena

Ribosomal RNA gene expression in a tetraploid H. bulbosum clone using biotin labelled in situ hybridization
Xu Jie, K.J. Kasha, and J.D. ProcunierDept. of Crop Science, University of Guelph
Guelph, Ontario, Canada. N1G 2WL
H. bulbosum has been used extensively in crosses with cultivated barley in order to either produce haploids for barley breeding (Kasha and Kao, 1970) or to transfer some useful variations such as disease resistance and winter hardiness into barley (Snape, et al., 1988). Based upon cytological observations, several researchers have suggested that suppression of nucleolus-organizing region (NOR) activity of H. bulbosum chromosomes in the interspecific hybrids could be related to chromosome elimination of H. bulbosum, giving rise to haploid production. Recently, Xu and Snape (1988) have confirmed that the H. bulbosum NOR is located on short arm of chromosome 6, based upon its homoeologous pairing with chromosome 6 of H. vulgare. They suggested that H. bulbosum chromosome 6 appears to be the first chromosome to be eliminated in the interspecific hybrids. Thus, it would be beneficial to obtain detailed information on NOR activity in H. bulbosum using in situ hybridization.
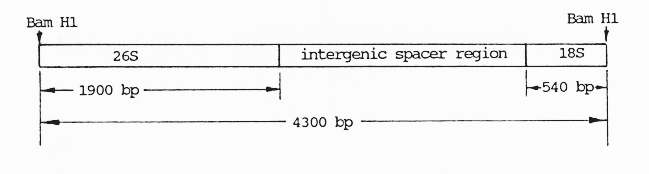
In this study, a tetraploid H. bulbosum clone (GBC141, 2n=4x=28) was used. The root tips were taken from plants and pretreated in ice- water for 10 hours before being fixed in acetic alcohol (1:3) for 24 hours. A 1% aceto-carmine solution was used to stain root tips before the preparations were made. pHbR26, a 4.3 kb region of the tandemly repeated ribosomal genes isolated from tetraploid H. bulbosum DNA by restriction with BAM H1 and cloned in the vector pBR322 (Kasha, unpublished), was used as the probe for in situ hybridization experiments (Fig. 1). The biotin labelling technique as described by Gustafson et al. (1988) was followed with the modification that incubation of preparations in a complex strapavidin-horseradish peroxidase (SA-HRP) and in diaminobenzidine tetrahydrochloride (DAB) was prolonged to 40-50 minutes and 3-4 hours, respectively.
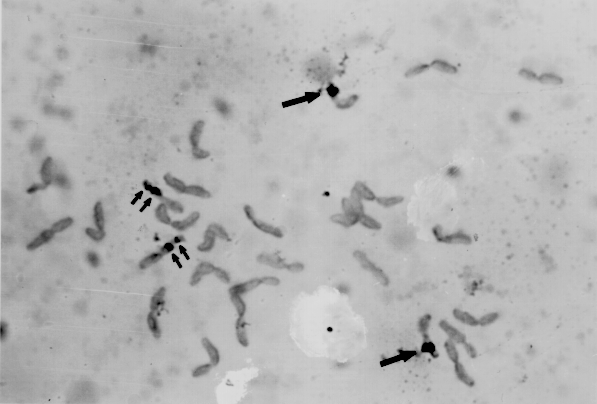
The hybridization sites with the biotinylated probe were detected clearly as dark spots against light green chromosome background (Fig. 2). Non-homologous hybridization sites, which occur invariably with radioactive labelling methods, were not observed. Therefore, the DNA sequence could be physically mapped on the homologous chromosome region with high precision. The increase of incubation time in SA-HRP and DAB solution tended to enhance the hybridization signal without causing background staining.
Tetraploid H. bulbosum has 28 chromosomes, four of which normally exhibit satellites in metaphase cells. We examined 20 metaphase cells stained with aceto-carmine for the secondary constrictions (gaps between the satellites and short arm of chromosome 6, Table 1). Although the number of visible gaps varied from I to 4, which was most likely due to the squashing procedures, most of cells (50%) showed 4 gaps as expected. This suggests that ribosomal genes in the 4 NORs are active.
Strong in situ hybridization signals were observed at the NOR region of all 4 satellite chromosomes as expected (Fig. 2). However, the observation that no more than 2 of the 4 chromosomes exhibited the gap in the biotin label of cells was unexpected. Ten cells with well- spread metaphase chromosomes were analyzed for a gap in the label (Table 1). In eight cells, only two of four satellite chromosomes showed clear gaps in the label. In the remaining two cells, only one of four satellite chromosomes exhibited the gap. Gustafson et al. (1988) interpreted the gaps expressed when using in situ hybridization as due to the active state of the ribosomal genes, whereas no gap meant an inactive state of the ribosomal genes, whereas no gap meant an inactive state in rye and wheat. This interpretation does not appear to be consistent with our observations. When stained with aceto-carmine, the gaps are present on all four of the NOR chromosomes of the H. bulbosum clone, indicating activity of rDNA genes. With in situ hybridization, no more than 2 of the 4 chromosomes exhibit a gap in the label. Rather than based upon activity or inactivity or NOR regions, we suggest an alternative hypothesis to explain the in situ observations. The NOR region consists of a large number (perhaps 2000-3000 in barley) of tandemly repeated genes for ribosomal RNA. Only a portion of these genes are normally expressed. If the portion being expressed which leads to the gap was in the middle of the tandemly repeated gene sequence, you would observe label on both sides of the gap. If the portion being expressed was a terminal section of the repeated sequence, you would not observe a gap in the in situ label. Our observations suggest that the regional expressed may be predetermined or possibly genetically controlled since the expression can be different for different chromosomes within the same plant.

It has been reported in cereals (Thompson and Flavell, 1988) that the ribosomal genes with a larger intergenic spacer region are preferentially transcribed. While such variation could also exist within a single plant of a cross-pollinated, self-incompatible species like H. bulbosum, the observation of gaps on all four chromosomes when stained with aceto-carmine makes this a less plausible explanation.
Acknowledgements:
The authors wish to express their thanks to Miss Rosalinda Oro for her kind assistance during the course of this study and to Miss Anne Dera for providing biotin labelling procedures and helpful discussions. Financial support from NSERC and OMAF are gratefully acknowledged.
References:
Gustafson, T.P., A.R. Dera, and S. Petrovic. 1988. Expression of modified rye ribosomal RNA gene in wheat. Proc. Natl. Acad. Sci. USA Vol. 85:3943-3945.
Kasha, K.J., and K.N. Kao. 1970. High frequency haploid production in barley (H. vulgare L.) Nature (Lond.) 225:874-876.
Snape, J.W., J. Xu, and B.B. Parker. 1988. Wide crossing and alien gene transfer. In Annual Report 1988, AFRC Institute of Plant Science Research and John Innes Institute. pp. 2-3.
Thompson, W.F., and R.B. Flavell. 1988. DNAse I sensitivity of ribosomal RNA genes in chromatin and nucleolar dominance in wheat. J. Mol. Biol. 204:535-548.
Xu, J., and J.W. Snape. 1988. The cytology of hybrids between H. vulgare and H. bulbosum revisited. Genome 30:486-494.
Xu, J., and J.W. Snape. 1989. The resistance of H. bulbosum and its hybrids with H. vulgare to common fungal pathogens. Euphytica (in press).